A new paper by Giuggioli et al. in PNAS September 30, 2013
Abstract:
Collective animal behavior studies have led the way in developing models that account for a large number of individuals, but mostly have considered situations in which alignment and attraction play a key role, such as in schooling and flocking. By quantifying how animals react to one another’s presence, when interaction is via conspecific avoidance rather than alignment or attraction, we present a mechanistic insight that enables us to link individual behavior and space use patterns. As animals respond to both current and past positions of their neighbors, the assumption that the relative location of individuals is statistically and history independent is not tenable, underscoring the limitations of traditional space use studies. We move beyond that assumption by constructing a framework to analyze spatial segregation of mobile animals when neighbor proximity may elicit a retreat, and by linking conspecific encounter rate to history-dependent avoidance behavior. Our approach rests on the knowledge that animals communicate by modifying the environment in which they live, providing a method to analyze social cohesion as stigmergy, a form of mediated animal–animal interaction. By considering a population of animals that mark the terrain as they move, we predict how the spatiotemporal patterns that emerge depend on the degree of stigmergy of the interaction processes. We find in particular that nonlocal decision rules may generate a nonmonotonic dependence of the animal encounter rate as a function of the tendency to retreat from locations recently visited by other conspecifics, which has fundamental implications for epidemic disease spread and animal sociality.
Conceptual Framework:
The delayed response between mark deposition, the action of an individual, and conspecific retreat, the reaction of another conspecific, is a basic ingredient of stigmergy (10, 11), a mediated interaction mechanism whereby the changes produced endogenously in the environment by the marks of one individual elicit a response in the neighbors, which in turn respond, affecting their nearest neighbors. This cascade of events creates a feedback mechanism for the entire population, which self-organizes into a dynamic spatiotemporal pattern.
The shrinking and growth of Si are controlled, respectively, by the aging of the marks and the movement of the animals. The transience of the deposited cues tends to reduce the size of a marked area, because inactive marks are ignored by conspecifics. An aging mark at a given location reduces the propensity of other conspecifics to retreat from that location, which in turn increases the pressure onto individual i to move further inside its own marked area, reducing even further the spatial extent of Si. Because a decrease in the size of Si further reduces its spatial extent, the decay of the marks acts as a positive feedback. The other positive feedback is the movement of the animals, which helps the growth of Si. As animals deposit marks by exploring regions beyond their inner-core areas, they increase the extent of Si and pressure neighbors into moving away to avoid confrontation. This in turn allows them to explore even larger areas, thus further increasing the size of their marked areas. Positive feedback mechanisms act to reinforce a given process and are the key to explaining various forms of aggregation and pattern formation (see, e.g., ref. 12 for the application of reinforced random walks to represent some types of positive feedback). On the other hand, the negative feedback acts in the opposite direction of the variation of Si whether it is a decrease or increase in its spatial extent. As marked areas get smaller, animals may traverse them quicker and thus slow their shrinking. Similarly, as marked areas get bigger, animals take longer to move across them, preventing individuals from re-marking aging marks. This results in a reduction of the growth rate of Si.
We choose to interpret the space use of marking animals as a stigmergic interaction for three reasons. The first is that animal marking is a widespread behavior in the animal kingdom, and although each species has evolved specialized means of communication by depositing cues on the terrain, it serves the general function of broadcasting an animal’s presence. Marks contain information about identity and relative dominance (13), with many vertebrates (14) and eusocial insects (15) making use of chemical signals but also with examples in which visual marks are used, such as feathers and feces by birds. Stigmergy represents a well-developed concept that would help in studying animal space use from a general theoretical perspective, independent of the types of signals present in the marks that get deposited or the sensory modalities required for the detection of those signals. The second reason is that stigmergy makes interactive processes history dependent, which captures the fact—often neglected in quantitative analyses of animal space use—that individuals do not respond simply to the current position of other conspecifics, but also to where they have been in the recent past. A mark, when detected, represents a record of an individual’s past activity in a specific location to which other conspecifics eventually respond. The third reason is that stigmergic stimulus–response association relies upon modification of the environment. As environmental heterogeneity may also affect how individuals move in space, our approach yields a method to quantify another form of spatial heterogeneity, the one generated endogenously from animal interactions. It thus may be possible to extend our current framework to provide a common currency to interpret animal space use as a function of the most important endogenous and exogenous features of the ecosystem, respectively, conspecific avoidance and environmental covariates. Promising approaches in that respect already are available and may help link population spatial distribution to animal spatial memory and landscape persistence (16), as well as to prey distribution and terrain steepness (17, 18).
In this framework of socially interacting animals, we are interested in determining how the individual movement response to the presence of conspecifics shapes the degree of segregation in the population. A useful tool to characterize the emerging spatiotemporal pattern of the population is the encounter rate of mobile animals, an instrument of broad ecological applicability (19). Most encounter estimates have relied upon basic animal movement models, in which displacement is ballistic and individuals are completely independent, which amounts to considering animals as “ideal gas” particles. This approach has been taken as a null model to estimate the frequency of meeting or associations among mobile animals (20) and has been used recently to estimate, with the help of allometric considerations in a spatially implicit context, how home range size scales with body mass (21). Here, to capture the key biological features of the movement and interaction processes that underlie animal spacing, we consider a spatially explicit scenario to determine how individual behavior affects animal space use. The focus of our analysis is the quantification of the average encounter rate, home range size, and degree of exclusivity as a function of the degree of stigmergy.
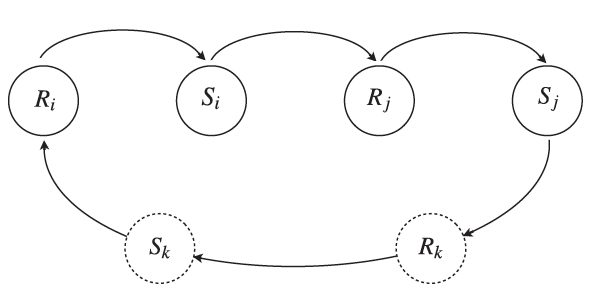
Fig. 1. Schematic representation of stigmergy in marking animals. When the animal i detects the presence of active foreign marks, it responds by retreating from the locations of the foreign marks (Ri). At the same time, the animal i itself deposits marks over the terrain, whose active set constitutes the stimulus Si that another member j of the population detects, inducing the response Rj. In turn, animal j deposits its own marks (Sj), whose locations affect animal i again or individual k, which will react and itself produce a stimulus. The number of individuals involved in this feedback loop may be as a large as the entire population or as small as just individuals i and j, depending on the locations the animals visit after their response. The dashed lines around Rk and Sk represent the fact that the number of steps necessary to affect individual i may vary because of the random nature of the movement process and, thus, of the probability of animal i encountering the stimulus (Sj).